11 வது தாவரவியல் : அலகு 11 : தாவரங்களில் கடத்து முறைகள்
தாவரங்களில் கடத்து முறைகள்
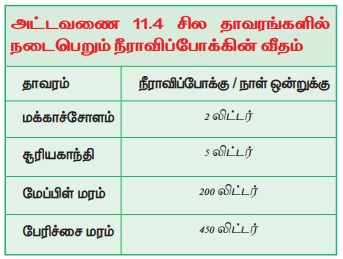
நீராவிப்போக்கு
வேர்களால் உறிஞ்சப்பட்ட நீர் இறுதியில் இலைகளை அடைந்து அங்கிருந்து நீராவியாக வளி மண்டலத்தினை அடைகிறது. உறிஞ்சப்பட்டதில் மிகச்சிறிய அளவிலான (5% ஐ விடக் குறைவு) நீரே தாவரத்தின் வளர்ச்சிக்கும் வளர்சிதை மாற்றத்திற்கும் பயன்படுத்தப்படுகிறது.
தாவரங்களின் பல்வேறு தரைமேல் பகுதிகளிலிருந்து அதிகப்படியான நீர், ஆவியாக வெளியேறுவதே நீராவிப்போக்கு எனப்படும். நீராவிப்போக்கு ஒருவகையான ஆவியாதல் என்றாலும் நீராவிப்போக்கு தாவரங்களுடன் தொடர்புடையது. நீராவிப்போக்கின் அளவு வியக்கத்தக்கது (அட்டவனை 11.4). நீராவிப்போக்கிற்காக சைலத்தின் வழியாக ஒரு நிமிடத்திற்கு 75 செ.மீ. என்ற வேகத்தில் நீரானது மேலேறிச் செல்கிறது.
செயல்பாடு
பள்ளி வளாகத்தில் இலைகளுடன் கூடிய கிளை உள்ள தாவரத்தினைத் தேர்வு
செய்து அக்கிளையினை ஒளிபுகும் பாலித்தின் பையால் மூடி கிளையின் அடிப்புறத்தில் கட்ட
வேண்டும். இரண்டு மணி நேரம் கழிந்த பின் நடைபெறும் மாற்றங்களை உற்று நோக்கி ஆசிரியரிடம்
விவாதிக்கவும்
1. நீராவிப்போக்கின் வகைகள் (Types of Transpiration)
கீழ்க்கண்ட மூன்று வகைகளில் நீராவிப்போக்கு நடைபெறுகிறது,
1. இலைத்துளை நீராவிப்போக்கு
இலையின் கீழ்ப்புறத் தோலில் அதிக அளவில் காணப்படும் நுண்ணிய துளைகளே இலைத்துளைகளாகும். தாவரங்களில் இலைத்துளை வழியாகவே மிக அதிக அளவில் (90 - 95%) நீராவிப்போக்கு நடைபெறுகிறது.
2. பட்டைத் துளை நீராவிப்போக்கு
தாவரங்களில் இரண்டாம் நிலை வளர்ச்சி நடைபெறுவதன் காரணமாகப் புறத்தோலுக்கு பதிலாக பெரிடெர்ம் உருவாகிறது. தண்டின் உள்ளமைந்த உயிருள்ள செல்களுக்கும் வெளிப்புற வளிமண்டலத்திற்கும் வாயுப்பரிமாற்றம் நடைபெறுவதற்கு ஏதுவாகச் சிறிய லென்ஸ் வடிவத் துளைகள் தண்டின் மேற்பரப்பில் காணப்படுகின்றன. இவையே லென்டிசெல் அல்லது பட்டைத் துளைகள் எனப்படும். மொத்த நீராவிப் போக்கின் அளவில் 0.1% மட்டுமே பட்டைத்துளை வழியாக நடைபெறுவதால் இது முக்கியத்துவமற்றதாகும்.
3. கியூட்டிகிள் நீராவிப்போக்கு
தாவரங்களின் இலைகள் மற்றும் தண்டின் புறத்தோலில் காணப்படும் கியூட்டின் எனும் கொழுப்பு பொருளால் ஆன மெழுகு அல்லது ரெசின் அடுக்கே கியூட்டிகிள் எனப்படும். கியூட்டிகிள் வழியாக மொத்த நீராவிப்போக்கில் மிகச்சிறிய அளவான 5 முதல் 10 சதவீதம் அளவிற்கே நீரிழப்பு நடைபெறுகிறது. வறண்ட நிலத் தாவரங்களில் கியூட்டிகிளின் தடிமன் அதிகரித்துக்காணப்படுவதால் நீராவிப்போக்கின் அளவு குறைந்தோ அல்லது அறவே இல்லாமலோ காணப்படுகிறது.
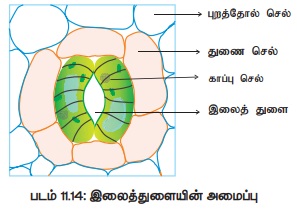
2. இலைத் துளையின் அமைப்பு.
இலைகளின் புறத்தோல் மற்றும் பசுமையான தண்டில் காணப்படும்
எண்ணற்ற சிறிய துளைகள் இலைத்துளைகள் எனப்படும்.
இலைத்துளையின் நீளம் மற்றும் அகலம் முறையே 10-40 மைக்ரான் மற்றும் 3-10 மைக்ரான் ஆகும்.
முதிர்ந்த இலைகளில் ஒரு சதுர மில்லி மீட்டருக்கு 50 முதல் 500 என்ற எண்ணிக்கையில் இவை
காணப்படும். இலைத்துளை இரண்டு காப்பு செல்களால் ஆனது. இவை பிறை நிலா அல்லது சிறுநீரக
வடிவத்தில் அமைந்த உயிருள்ள புறத்தோல் செல்களாகும். காப்பு செல்களைச் சுற்றிக் காணப்படும்
புறத்தோல் செல்களுக்குத் துணை செல்கள்
என்று பெயர். காப்பு செல்களின் இரு முனைகள் இணைந்தும் முனைகளுக்கு இடைப்பட்ட பகுதி
இணையாமலும் இருப்பதால் துளை வடிவம் உருவாகிறது. காப்பு செல்லின் உட்புறச் சுவர் வெளிப்புறச்
சுவரை விட தடித்தது (படம் 11.14). இலைத்துளையின் உட்புறம் காணப்படும் குழி இலைத்துளை அறை எனப்படும். இது செல்லிடைவெளிகளுடன்
தொடர்பு கொண்டது.
3. இலைத்துளை இயக்கத்தின் செயல்முறைகள்:
காப்பு செல்களில் ஏற்படும் விறைப்பழுத்தமே இலைத்துளை
இயக்கத்தினைக் கட்டுப்படுத்துகிறது. காப்பு செல்களுக்குள் நீர் செல்லும்போது சீரற்ற
சுவர் தடிப்பு காரணமாக விரிவடைந்து இலைத்துளை திறக்கிறது. காப்பு செல்லின் வெளிச்சுவர்
குவித்தும் இழுபடும் தன்மை கொண்டு இருப்பதால் அது இழுபடு தன்மையற்ற உட்புறச் சுவரினை
வெளி நோக்கி இழுப்பதன் மூலம் இலைத் துளை திறக்கிறது.
பல்வேறு வகையான கோட்பாடுகள் இலைத்துளை திறந்து மூடுதலை
விவரிக்கின்றன. அவற்றுள் முக்கியமான கோட்பாடுகள் பின்வருமாறு,
1. காப்பு செல்லின் ஒளிச்சேர்க்கை கோட்பாடு
2. தரச- சர்க்கரை இடைமாற்ற கோட்பாடு
3. பொட்டாசியம் அயனியின் செயலூக்கக் கடத்தல் கோட்பாடு
1. காப்பு செல்லின் ஒளிச்சேர்க்கை கோட்பாடு
இலைத்துளைகள் பகலில் திறந்திருப்பதையும் இரவில் மூடுவதையும் ஃபான் மோல் (1856) கண்டறிந்தார். இவரின் கூற்றுப்படி காப்பு செல்லில் உள்ள பசுங்கணிகங்கள் பகலில் ஒளிச்சேர்க்கை செய்வதால் சர்க்கரை உற்பத்தி செய்கின்றது. இது காப்பு செல்லின் சவ்வூடுபரவல் அழுத்தத்தினை அதிகரித்து அருகமைந்த செல்களில் இருந்து நீர் உட்புகுந்து இலைத் துளையினைத் திறக்கிறது. இரவில் இதற்கு எதிரான செயல் நடைபெறுவதால் இலைத்துளை மூடுகிறது.
குறைகள் :
அ) காப்பு செல்லில் உள்ள பசுங்கணிகங்கள் முழுமையான வளர்ச்சி அடையாததால் இவற்றால் ஒளிச்சேர்க்கை செய்ய இயலாது.
ஆ) காப்பு செல்லில் இயல்பாகவே சர்க்கரை அதிக அளவு காணப்படும்
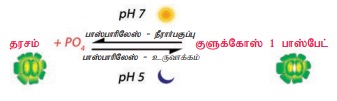
2. தரச - சர்க்கரை இடைமாற்ற கோட்பாடு
அ) லாயிட் (1908) என்பாரின் கூற்றுப்படி, காப்பு செல்லின் விறைப்புத் தன்மை என்பது தரச - சர்க்கரை இடைமாற்றத்தினை சார்ந்தது. இக்கருத்தினை ஆதரித்த லாப்ட்ஃபீல்ட் (1921) காப்பு செல்கள் பகலில் திறந்திருக்கும்போது சர்க்கரையும் இரவில் மூடியிருக்கும்போது தரசம் கொண்டிருப்பதையும் கண்டார்.
ஆ) ஸேயேர் (1920), இலைத்துளைகள் திறந்து மூடுவதற்கு காப்பு செல்களில் ஏற்படும் அமில-கார (pH) வேறுபாடே காரணம் எனக் கண்டறிந்தார். இவரின் கூற்றுப்படி பகலில் அதிக pH இருப்பதால் இலைத்துளை திறந்தும் இரவில் குறைந்த pH இருப்பதால் இலைத்துளை மூடியும் காணப்படுகிறது. பகலில் கார்பன்டை ஆக்சைடானது ஒளிச்சேர்க்கையில் பயன்படுத்தப்படுவதால் pH அளவு உயர்ந்து தரசம் சர்க்கரையாக மாறுகிறது. இச்சர்க்கரையின் அதிகரிப்பு உட்சவ்வூடுபரவலை ஊக்குவித்து விறைப்பழுத்தத்தினை அதிகரிப்பதால் இலைத்துளை திறக்கிறது. இதேபோன்று இரவில் கார்பன் டை ஆக்ஸைடு செல்களில் அதிகரிப்பதால் pH அளவு குறைந்து சர்க்கரை தரசமாக மாற்றமடைகிறது. தரசம் காப்பு செல்லின் விறைப்பழுத்தத்தினைக் குறைப்பதால் இலைத்துளை மூடுகிறது.
இ) தரச-சர்க்கரை கோட்பாட்டிற்கு வலு சேர்க்கும் வகையில்
ஹேன்ஸ்
(1940) என்பவரால் காப்பு செல்லில் பாஸ்பாரிலேஸ் நொதி கண்டறியப்பட்டது. பகலில்
பாஸ்பாரிலேஸ் நொதி தரசத்தினை நீராற்பகுத்து சர்க்கரையாக மாற்றி pH- அளவை உயர்த்துவதால்
உட்சவ்வூடுபரவல் நடைபெற்று இலைத்துளை திறக்கிறது. இரவில் இதற்கு எதிரான செயல் நடைபெறுகிறது.
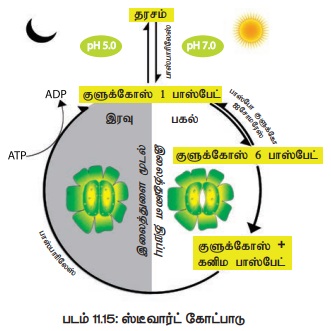
ஈ) ஸ்டீவார்ட் (1964) சற்று மாற்றியமைக்கப்பட்ட தரச- சர்க்கரை இடைமாற்ற கோட்பாட்டினை முன்மொழிந்தார். இதன்படி குளுக்கோஸ் -1- பாஸ்பேட் சவ்வூடுபரவல் தன்மையற்றது. குளுக்கோஸ்-1- பாஸ்பேட்டில் உள்ள பாஸ்பேட்டை நீக்குவதால் அது குளுக்கோஸாக மாறுகிறது. குளுக்கோஸின் சவ்வூடுபரவல் திறன் தன்மையால் காப்பு செல்லினுள் இதன் செறிவு அதிகரித்து இலைத்துளை திறக்கிறது (படம் 11.15).
தரச - சர்க்கரை இடைமாற்ற கோட்பாட்டிற்கான எதிர்ப்புகள் :
1) ஒரு விதையிலைத் தாவரத்தின் காப்பு செல்லில் தரசம்
இல்லை.
2) இலைத்துளை திறக்கும்போது சர்க்கரை அதிகரித்துத்
தரசம் குறைவதற்கான ஆதாரம் இல்லை.
3) pH -ன் அளவானது 5-லிருந்து 7 ஆக மாறுவதற்குக் காரணம் கார்பன் டை ஆக்ஸைடில் ஏற்படும் மாற்றங்களால் தான் என்பதை இக்கோட்பாட்டால் விளக்க இயலவில்லை.
3. பொட்டாசியம் அயனி கடத்தல் கோட்பாடு
லெவிட் (1974) என்பவரால் வெளியிடப்பட்ட இக்கொள்கையினை ராஷ்க் (1975) விளக்கினார். இதன்படி இலைத் துளை திறத்தல் கீழ்க்கண்ட படி நிலைகளை உள்ளடக்கியது.
பகலில் :
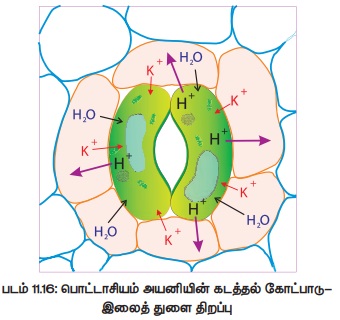
அ) காப்பு செல்லில் தரசம் கரிம அமிலமாக (மாலிக் அமிலம்) மாற்றமடைகிறது.
ஆ) காப்பு செல்லில் உள்ள மாலிக் அமிலம் மாலேட் எதிர்மின் அயனியாகவும் புரோட்டானாகவும் (H+) பிரிகிறது.
இ) புரோட்டான்கள் சவ்வின் வழியாக அருகமைந்த துணை செல்களுக்கும், துணை செல்களிலிருந்து பொட்டாசியம் அயனிகள் (K+) காப்பு செல்களுக்கும் பரிமாற்றம் செய்து கொள்கின்றன. இது மின் சரிவு வாட்டத்தின் வழியாக (Electrical Gradient) நடைபெறுவதால் இது அயனிப்பரிமாற்றம் (lon Exchange) எனப்படுகிறது.
ஈ) அயனிப்பரிமாற்றம் ஆற்றல் தேவைப்படும் செயலாதலால் இதற்கு ATP பயன்படுத்தப்படுகிறது.
உ) காப்பு செல்லில் பொட்டாசியம் அயனி மிகுவதால் அதனைச் சமன்படுத்தக் குளோரைடு (CI-) அயனி பயன்படுத்தப்படுகிறது. இதனால் காப்பு செல்லில் கரைபொருள் அடர்த்தி அதிகரித்து நீரியல் திறன் குறைகிறது.
ஊ) இதன் விளைவாகக் காப்பு செல் ஹைப்பர்டானிக் நிலையினை அடைந்து அருகில் அமைந்த செல்களில் இருந்து நீர் உள்ளே நுழைகிறது...
எ) நீர் உட்புகுவதால் விறைப்பழுத்தம் அதிகரித்து இலைத்துளை திறக்கிறது (படம் 11.16)
இரவில் :
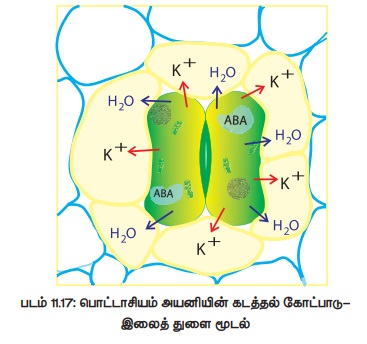
அ) இரவில் ஒளிச்சேர்க்கை நடைபெறுவதில்லை என்றாலும் சுவாசித்தல் தொடர்ந்து நடைபெறுவதால் இலைத்துளை அறையில் கார்பன்-டை-ஆக்ஸைடு சேகரமாகிறது.
ஆ) இவ்வாறு சேகரமான கார்பன்-டை-ஆக்ஸைடு செல்லுக்குள் pH அளவினைக் குறைக்கிறது.
இ) காப்பு செல்லில் ஏற்படும் குறைவான pH -ம் நீர் பற்றாக்குறையும் நெருக்கடி நிலை ஹார்மோனான அப்சிசிக் அமிலத்தினை (ABA) தூண்டுகிறது.
ஈ) அப்சிசிக் அமிலம் துணை செல்களிலிருந்து உள்வரும் பொட்டாசியம் அயனியினைத் தடுத்தும் முன்பே காப்பு செல்களில் உள்ள பொட்டாசியம் அயனியினை வெளியேற்றவும் செய்கிறது.
உ) இதனால் காப்பு செல்களில் நீரிழப்பு ஏற்பட்டு அதன் விறைப்பழுத்தத்தினை குறைப்பதால் இலைத்துளை மூடுகிறது (படம் 11.17).
4. நீராவிப்போக்கின் வீதத்தை பாதிக்கும் காரணிகள்
நீராவிப்போக்கின் வீதத்தைப் பாதிக்கும் காரணிகள் இரண்டு
வகைகளாகப் பிரிக்கப்பட்டுள்ளன. அவை
1) வெளிப்புறக் காரணிகள் அல்லது சுற்றுச் சூழல் காரணிகள்
2) உட்புறக் காரணிகள் அல்லது தாவரக் காரணிகள்
1) வெளிப்புறக் காரணிகள் அல்லது சுற்றுச் சூழல் காரணிகள்
அ) வளிமண்டல ஈரப்பதம் : வளிமண்டத்தில் மிகுந்த ஈரப்பதம் இருக்கும்போது நீராவிப்போக்கின் வீதம் வெகுவாகக் குறைகிறது. உலர்ந்த காற்று நீராவிப்போக்கின் வீதத்தை நேர்தகவில் அதிகரிக்கிறது.
ஆ) வெப்பநிலை: வளிமண்டலத்தின் வெப்பநிலை அதிகரிக்கும்போது நீராவிப்போக்கின் வீதமும் அதிகரிக்கிறது. ஆனால் அதிகப்படியான வெப்பநிலையில் இலைத்துளையின் விறைப்பழுத்தம் குறைவதால் இலைத்துளை மூடப்படுகிறது.
இ) ஒளி: ஒளியின் செறிவு வெப்பநிலையை அதிகரிக்கிறது. வெப்பநிலையினைப் போலவே அதிக ஒளிச் செறிவில் நீராவிப்போக்கு அதிகரித்தும் குறைந்த ஒளிச்செறிவில் நீராவிப்போக்கு குறைந்தும் காணப்படுகிறது. அதிகப்படியான ஒளி செல் சவ்வின் கடத்துத்தன்மையினை அதிகரித்து நீர் மூலக்கூறுகள் செல்லை விட்டு வெளியேறுவதை எளிதாக்குகிறது.
ஈ) காற்றின் திசைவேகம் : காற்று வீசாத நிலையில் இலைத்துளைப் பரப்பிற்கு மேல் நீர் மூலக்கூறுகள் பூரிதத் தன்மையினை அடைந்துள்ளதால் அதிகப்படியான நீர் ஆவியாவதில்லை. காற்று வீசும்போது, இலைப்பரப்பிற்கு மேலுள்ள நீராவிப்படலம் அகற்றப்படுவதால் பரவல் அழுத்தப் பற்றாக்குறை ஏற்பட்டு இலையின் செல்களில் இருந்து நீர் ஆவியாகி நீராவிப்போக்கு அதிகரிக்கிறது. ஆனால் அதிக திசைவேகம் கொண்ட காற்று அதிகப்படியான நீரிழப்பினை ஏற்படுத்தி இலைத்துளைமூடப்படுகிறது. இதனால் நீராவிப்போக்கின் வீதம் குறைகிறது.
செயல்பாடு: ஒரு தாவரத்தினை மின் விசிறியின் கீழும் மற்றும் குளிர்சாதன அறையிலும் வைத்தால் என்ன நிகழும்?
உ) வளிமண்டல அழுத்தம்: குறைந்த வளி மண்டல அழுத்தத்தில் நீராவிப்போக்கின் வீதம் அதிகரிக்கிறது. மலைப்பகுதியில் குறைந்த வளி மண்டல அழுத்தம் நிலவுவதால் அங்கு நீராவிப்போக்கின் வீதம் அதிகமாக இருக்க வேண்டும். ஆனால் அங்கு நிலவும் குறைந்த வெப்பநிலை இதனை ஈடுசெய்வதால் நீராவிப்போக்கின் வீதம் கட்டுக்குள் வைக்கப்படுகிறது.
ஊ) நீர் : தாவரத்தின் இயல்பான வளர்ச்சிக்கு மண்ணில் போதிய அளவு நீர் இருக்க வேண்டும். நீராவிப்போக்கின் காரணமாக ஏற்படும் அதிகப்படியான நீரிழப்பு தாவரங்களுக்கு வாடலை (Wilting) ஏற்படுத்துகிறது. பொதுவாக வாடல் முன்று வகைப்படும் அவை,
(1) தொடக்க நிலை வாடல் : தாவரச் செல்லில் நீரின் அளவு குறைகிறது. இவ்வகையில் அறிகுறிகள் வெளிப்படையாகத் தெரிவதில்லை
(2) தற்காலிக வாடல் : கடும் கோடையின் பகல் பொழுதில் இளந்தாவரங்களில் ஏற்படும் நீரிழப்பு வாடலை ஏற்படுத்தினாலும் இரவுப் பொழுதில் அவை மீள் நிலையினை அடைகின்றன.
(3) நிரந்தர வாடல்
: நீர் உறிஞ்சப்படுதல் நிறுத்தப்படுவதால் தாவரச் செல்களுக்கு
நீர் கிடைக்காமல் நிரந்தர வாடல் நிலைக்கு இட்டுசெல்கிறது.
2) உட்புறக் காரணிகள் அல்லது தாவரக் காரணிகள்
அ) இலைப் பரப்பு: இலைப்பரப்பின் அதிகரிப்பு நீராவிப்போக்கினையும் அதிகரிக்கும்.
எனவே வறண்ட நிலத்தாவரங்களில் இலைப்பரப்பு குறைந்து காணப்படுகிறது.
ஆ) இலையின் அமைப்பு: நீராவிப்போக்கினைக் குறைப்பதற்காக இலையில் புதையுண்ட
இலைத்துளைகள், தூவிகள், கியூட்டிகிள், நீர்விரும்பும் பொருட்களான பசை, மியூசிலேஜ் ஆகிய
உள்ளமைப்புகள் காணப்படுகின்றன. இத்தகைய மாற்றங்கள் வறண்ட நிலத் தாவரங்களில் குறிப்பிடத்
தகுந்த அளவில் காணப்படுகின்றன. ஒபன்ஷியாவில் நீராவிப்போக்கை குறைப்பதற்காக இலைபோன்று
அமைந்த தண்டு காணப்படுகிறது. இதற்குப் பில்லோகிளாடு என்று பெயர். ஆஸ்பராகஸ் தாவரத்தில் வரம்புடைய வளர்ச்சியுடைய
தண்டு இலைகள் போன்ற அமைப்புகளில் காணப்படுகிறது. இது கிளாடோடு அல்லது கிளாடோபில்
எனப்படும். மேலும் சில தாவரங்களில் இலைக்காம்பு தட்டையாகி அகன்று காணப்படும் இது
பில்லோடு எனப்படும். எடுத்துக்காட்டு: அகேஷியா மெலனோசைலான்.
5. தாவர நீராவிப்போக்குத் தடுப்பான்கள் (Plant Antitranspirants)
தாவரத்தில் நீராவிப்போக்கினைத் தடுக்கப் பயன்படும் எந்தப் பொருளும் நீராவிப்போக்குத் தடுப்பான்கள் எனப்படும். ஒரு சிறந்தநீராவிப்போக்குத் தடுப்பான் தாவரத்தின் வாயுப்பரிமாற்றத்திற்கு எவ்வித ஊறும் விளைவிக்காமல் நீராவிப்போக்கினை தடுக்க வேண்டும். தாவர நீராவிப்போக்குத் தடுப்பான்கள் இரு வகைப்படும்
1) இலைத்துளைக்கு
மேல் இயல்சார் தடுப்பான்களாகச் செயல்படுபவை
நிறமற்ற பிளாஸ்டிக், சிலிக்கோன் எண்ணெய் மற்றும் குறைந்த பாகுத்தன்மை மெழுகுகள் ஆகியவற்றை மெல்லிய படலமாக இலையின் மேல் தெளிப்பதால் நீராவிப்போக்கு தடைபடுகிறது. ஆனால் இப்படலம் கார்பன் டை ஆக்ஸைடு மற்றும் ஆக்ஸிஜன் ஆகியவற்றின் ஊடுருவலை தடை செய்வதில்லை. இயல்சார் தடுப்பான்களின் வெற்றி மிகவும் குறைவான சதவீதமேயாகும்.
2) இலைத்துளை
மூடுவதை தூண்டுபவை
கார்பன்-டை-ஆக்ஸைடு இலைத்துளை மூடலைத் தூண்டுவதால் இது ஒரு இயற்கையான நீராவிப்போக்குத் தடுப்பான் ஆகும். மேலும் இதனைப் பயன்படுத்துவதால் ஒளிச்சுவாசமும் தடைபடுவது கூடுதலான நன்மையாகும். பினைல் மெர்குரி அசிடேட்டினை இலையில் தெளிக்கும் போது அது இரண்டு வார காலம் அல்லது கூடுதல் காலத்திற்கு எவ்வித நச்சு பின்விளைவின்றி இலைத்துளையினைப் பகுதி அளவில் மூட வைக்கிறது. அப்சிசிக் அமிலம் இலைத்துளை மூடுதலைப் பெருமளவில் தூண்டுகிறது. டோடிசினில் சக்சினிக் அமிலமும் இலைத் துளை மூடுதலில் பங்கு வகிக்கிறது.
பயன்கள் :
• நீராவிப்போக்குத் தடுப்பான்கள் பயிர் தாவரங்களில் ஏற்படும் பெருமளவு நீர் இழப்பினைத் தடுக்கிறது.
• நாற்றுப்பண்ணைகளில் மறுநடவு செய்யப் பயன்படுகின்றன.
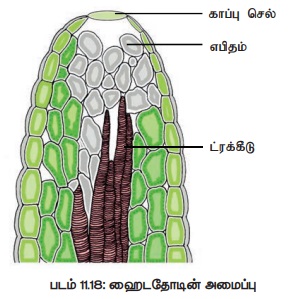
6. நீர் வடிதல் (Guttation)
காற்றில் ஈரப்பதம் அதிகமாக இருக்கும் போது நீராவிப்போக்கின்
வீதம் வெகுவாகக் குறைகிறது. இந்நிலையில் நீர் உறிஞ்சப்படும் போது தாவரத்தினுள் அதிகப்படியான
நீர் சேர்ந்து வேர் அழுத்தத்தினை உருவாக்குகிறது. இந்த அதிகப்படியான நீர் தாவர இலைகளின்
விளிம்புகளில் நீராக வடிகிறது. இதுவே நீர்
வடிதல் எனப்படுகிறது. எடுத்துக்காட்டு: புற்கள், தக்காளி, உருளைக்கிழங்கு, கத்திரி
மற்றும் அலோகேஷியா. ஈரப்பதமுள்ள நிழற்பாங்கான
பகுதியில் வளரும் தாவரங்களில் காணப்படும் இவ்வகை நீர்வடிதலில் இலைத்துளை போன்ற ஒரு
துளை வழியாகவே நீர் வெளியேறுகிறது. இத்துளை நீர்சுரப்பி
அல்லது ஹைடதோடு எனப்படும். இத்துளையினுள் அதிகச் செல்லிடைவெளி கொண்டு பரவலாக
அமைந்த செல்கள் காணப்படுகின்றன. இவை எபிதெம் எனப்படும் (படம் 11.18). எபிதெம்,
நரம்புகளின் சைலம் மற்றும் ஃபுளேயம் முடிவில் காணப்படும். ஹைடதோடுகள் வழியாக வரும்
நீர் தூய்மையானதல்ல. இது பல்வேறு கரைபொருட்களைப் பெற்ற கரைசலாகும்.
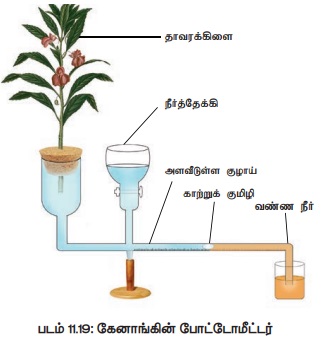
7. நீராவிப்போக்கினை அளவிடுதல்
1) கேனாங்கின் போட்டோமீட்டர்
கேனாங்கின் போட்டோமீட்டர் நீராவிப்போக்கினை மறைமுகமாக அளவீடு செய்யப் பயன்படுகிறது. இம்முறையில் உறிஞ்சப்படும் நீரின் அளவினை அளப்பதன் மூலமாக ஆவியாகும் நீராவிப்போக்கின் அளவு அனுமானிக்கப்படுகிறது.
ஒரு கிடைமட்டமான குறுகிய குழாயின் இரு முனைகளும் ஒன்றுக்கொன்று
எதிராகவளைந்ததாக இக்கருவி அமைந்துள்ளது. ஒருபுறம் உள்ள வளைந்த முனை அகன்றதாகவும் மறுபுறம்
உள்ள வளைந்த முனை குறுகியதாகவும் உள்ளது. இத்துடன் திருகு மறையுடன் கூடிய நீர்தேக்கியானது
அகலமான முனைக்கு அருகில் உள்ளது. அகலமான முனையில் ஒரு தாவரக் கிளை அல்லது சிறு தாவரத்தினை
பிளவுற்ற ரப்பர் அடைப்பானின் உதவியால் பொருத்தப்பட்டுள்ளது. கிடைமட்டக் குழாயின் மறுமுனை
வண்ண நீர் நிரம்பிய பீக்கருள் மூழ்கியவாறு உள்ளது (படம் 11.19). இக்குறுகிய முனையினுள்
காற்றுக் குமிழி தோற்றுவிக்கப்பட்டு இந்த அமைப்பை சூரிய ஒளியில் வைத்து உற்று நோக்க
வேண்டும் நீராவிப்போக்கு நடைபெறுவதால் காற்றுக்குமிழ் அகலமான முனையினை நோக்கிச் செல்கிறது.
நீராவிப் போக்கினால் ஏற்படும் நீர் இழப்பை ஈடுகட்ட தாவரக் கிளையின் சைலம் மூலம் நீர்
உறிஞ்சப்படுகிறது. எனவே நீராவிப்போக்கின் வீதமும் நீர் உறிஞ்சுதல் வீதமும் சமமானது
என அறியப்படுகிறது.
2. கோபால்ட் குளோரைடு (CoCl2) தாள் முறை
பசுமையான மேல்கீழ் வேறுபாடுடைய இலை ஒன்றினைத் தேர்வு
செய்து அதன் இரு புறத்தினையும் உலர்ந்த பஞ்சியினைக் கொண்டு சுத்தம் செய்ய வேண்டும்.
இலையின் இரு புறத்தையும் உலர்ந்த கோபால்ட் குளோரைடு காகிதத் துண்டினால் மூடி அதன் மேல்க
ண்ணாடித் துண்டுகளை அசையாதவாறு பொருத்த வேண்டும். இந்த அமைப்பினை சிறிது நேரம் இடையூறின்றி
வைக்கும்போது இலையின் அடிப்புறத்தோலில் உள்ள கோபால்ட் குளோரைடு தாள் இளஞ்சிவப்பு நிறமாக
மாறியுள்ளதைக் காணலாம். இதற்குக் காரணம் இலைத்துளை வழியாக வரும் நீராவியினால் கோபால்ட்
குளோரைடு தாள் நீரேற்றம் (CoCl2
2H20 அல்லது Cocl2.4H2O)
பெறுவதேயாகும். மேலும் இவ்வாறான மேல்கீழ் வேறுபாடடைந்த இலையில் நீராவிப்போக்கின் வீதம்
மேற்புறத்தோலை விட அடி புறத்தோலில் அதிகம் இருப்பதையும் அறியலாம்.
8. நீராவிப்போக்கின் முக்கியத்துவம்
நீராவிப்போக்கு நீரிழப்பிற்கு வழிகோலுகிறது. இப்பாடத்தின் முன்பகுதியில் குறிப்பிட்டதுபோல உள்ளெடுக்கப்பட்ட நீரில் 95 விழுக்காடு நீராவிப்போக்கினால் இழக்கப்படுகிறது. பொதுவாக இது தாவரத்திற்குக் கேடு தரும் செயல்பாடு எனக் கருதக்கூடும். ஆனால் நீரின் உள்ளெடுப்பு, சாறேற்றம், கனிம உள்ளெடுப்பு ஆகியவை நேரடியாக நீராவிப்போக்கினை சார்ந்துள்ளன. இன்னும் கூறப்போனால் தாவரங்கள் சுட்டெரிக்கும் சூரிய ஒளியில் பசுமையுடன் இருப்பதற்குக் காரணம் நீராவிப்போக்கே. எனவே கர்டிஸ் கூற்றுப்படி நீராவிபோக்கு என்பது ஒரு "தேவையான தீமை" ஆகும்.