தாவரவியல் - கனிமங்களின் உள்ளெடுப்பு | 11th Botany : Chapter 11 : Transport in Plants
11 வது தாவரவியல் : அலகு 11 : தாவரங்களில் கடத்து முறைகள்
கனிமங்களின் உள்ளெடுப்பு
கனிமங்களின் உள்ளெடுப்பு (Mineral Absorption)
கனிமங்கள் நிலக்கரைசலில் கரைநிலை அல்லது மண்ணின் கூழ்மத்துக்களுடன் இணைந்த நிலை என இரு வகையான அமைப்பில் காணப்படுகின்றன. கனிமங்களின் உள்ளெடுப்பு நீரின் உள்ளெடுப்புடன் சேர்ந்தே நடைபெறுகிறது என முற்காலத்தில் தவறுதலாகக் கருதப்பட்டது. ஆனால் கனிமங்களின் உள்ளெடுப்பும் நீரின் உள்ளெடுப்பும் ஒன்றையொன்று சாராத இரு தனித்தனிச் செயல்களாக உள்ளன. கனிமங்கள் வேர்த்தூவிகளால் மட்டும் உறிஞ்சப்படுவதில்லை, வேரின் புறத்தோல் அடுக்காலும் (எபிபிளம்மா) உள்ளெடுக்கப்படுகிறது.
வேர் செல்லின் பிளாஸ்மா சவ்வு தன்னைக் கடந்து செல்ல அனைத்து அயனிகளையும் அனுமதிப்பதில்லை. அதேபோல, ஒரே கனிம உப்பில் உள்ள அனைத்து அயனிகளும் ஒரே சமமான அளவில் உள்ளெடுக்கப்படுவது இல்லை.
உயிருள்ள செல்களுக்குள் அல்லது திசுக்களுக்குள் அவற்றைச் சூழ்ந்துள்ள ஊடகத்திலிருந்து செல் சவ்வினைத் தாண்டி அயனிகள் ஊடுருவுதலும், சேகரமாதலும் கனிம உள்ளெடுப்பு எனப்படும். செல் அல்லது திசுவிற்கு உள்ளேயும் வெளியேயும் நடைபெறும் அயனிகளின் இடப்பெயர்விற்கு அயனிகடத்தல் அல்லது அயனிபுகல் (Flux) என்று அழைக்கப்படுகிறது. அயனிகள் செல்லுக்குள் செல்வதை அயனி உட்புகல் (Influx) என்றும் செல்லிலிருந்து வெளியேறுதல் அயனி வெளிப்புகல் (Efflux) என்றும் அழைக்கப்படும். கனிம உள்ளெடுப்பின் செயல்முறைகளை விளக்குவதற்கு எண்ணற்ற கோட்பாடுகள் உள்ளன. இக்கோட்பாடுகளை ஆற்றல் சாரா செயல்முறை (வளர்சிதை மாற்றச் சக்தி தேவைப்படாதது) மற்றும் ஆற்றல் சார் செயல்முறை (வளர்சிதை மாற்றச் சக்தி தேவைப்படுவது) என வகைப்படுத்தலாம்.
1. ஆற்றல்சாரா உள்ளெடுப்பு
1. அயனிப் பரிமாற்றம் (Ion-Exchange)
வேர் செல்களுக்குள் இருக்கும் அயனியும் நிலக்கரைசலில்
அதே மின்சுமையுடன் உள்ள அயனியும் பரிமாறிக்கொள்ளப்படுவது அயனிப் பரிமாற்றமாகும். இதனை
இரு கோட்பாடுகள் மூலம் விளக்கலாம். அவை i. தொடர்பு பரிமாற்றக் கோட்பாடு மற்றும்
ii. கார்பானிக் அமில பரிமாற்றக் கோட்பாடு.
அ. தொடர்பு பரிமாற்றக்
கோட்பாடு (Contact Exchange Theory)
இக்கொள்கையின்படி வேர்செல்களின் மேற் பரப்பில் உள்ள
அயனிகளும் மண் துகளில் ஒட்டியுள்ள அயனிகளும் உறுதியாகப் பிணையாமல் ஒரு குறுகிய வட்டப்பரப்பில்
ஊசலாடிக் கொண்டுள்ளன. இப்பரப்பு ஊசலாடும் பரப்பு
(Oscillation volume) எனப்படும். இக்குறுகிய
பரப்பு காரணமாக அயனிகள் ஒன்றுடன் ஒன்று மோதுவதால் தங்களுக்குள் எளிதாகப் பரிமாற்றம்
செய்து கொள்கின்றன. (படம் 11.23)
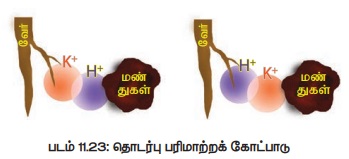
ஆ. கார்பானிக் அமிலப் பரிமாற்றக் கோட்பாடு (Carbonic Acid Exchange Theory)
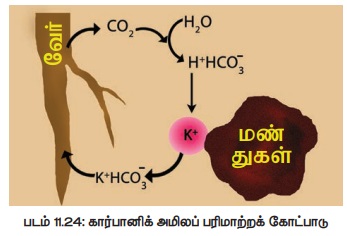
இக்கொள்கையின்படி, அயனிப்பரிமாற்றத்திற்கு மிக முக்கிய பங்கு வகிப்பது நிலக் கரைசலாகும். இதுவே அயனிப்பரிமாற்றத்திற்கு ஊடகமாகவும் செயல்படுகிறது. வேர் செல்களில் நடைபெறும் சுவாசித்தல் காரணமாக வெளிப்படும் கார்பன் டை ஆக்ஸைடானது நீருடன் சேர்ந்து கார்பானிக் அமிலமாக (H2CO3) மாறுகிறது. இவ்வமிலம் மண் கரைசலில், புரோட்டானாகவும் (H+) பைகார்பனேட்டாகவும் (HCO3- ) பிரிகிறது. இதில் புரோட்டான்கள் மண் துகளில் பிணைந்துள்ள நேர்மின் அயனியுடன் பரிமாறிக் கொள்ளப்படுவதால் நேர்மின் அயனியானது மண் துகளிலிருந்து விடுவிக்கப்பட்டு மண் கரைசலில் சேர்கிறது. பின் அது எளிதில் வேர்செல்களால் உள்ளெடுக்கப்படுகிறது (படம் 11.24).
2. ஆற்றல்சார் உள்ளெடுப்பு
செறிவு சரிவு வாட்டத்திற்கு எதிராக வளர்சிதை மாற்ற ஆற்றலினைப் பயன்படுத்தி நடைபெறும் அயனிகளின் உள்ளெடுப்பு ஆற்றல்சார் உள்ளெடுப்பு எனப்படும். தாவரங்களில் உள்ள வாக்குவோல்களில் நேர்மின் அயனிகளும் எதிர்மின் அயனிகளும் செறிவு சரிவு வாட்டத்திற்கு எதிராகச் சேகரமாவதை ஆற்றல் சாரா உள்ளெடுப்பால் விளக்கமுடியவில்லை. தாங்கிக் கடத்திக் கருத்துரு வழியாகக் கனிமங்களின் ஆற்றல்சார் உள்ளெடுப்பினை விளக்கலாம்.
தாங்கிக் கடத்திக் கருத்துரு (The Carrier Concept)
ஃபான் டென் ஹானெர்ட் என்பவரால் 1937ஆம் ஆண்டு இக்கருத்துரு முன்மொழியப்பட்டது.
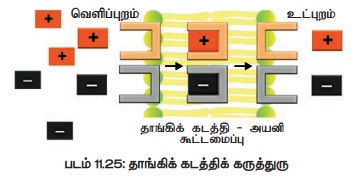
இதன்படி செல்சவ்வானது அயனிகளைத் தாமாக உள் ஊடுருவுவதை அனுமதிப்பதில்லை. எனவே செல் சவ்வில்
அமைந்துள்ள கடத்தி மூலக்கூறுகள் ஒரு ஊர்தி போலச் செயல்பட்டு அயனிகளை சுமந்து கொண்டு
கடத்தி-அயனி கூட்டமைவாக உருமாறிச் செல்சவ்வினைக் கடக்கின்றன. இந்த கடத்தி - அயனி கூட்டமைவானது
செல்லின் உட்பரப்பில் பிரிந்து அயனிகளை விடுவித்த பிறகு கடத்தியானது மீளவும் புதிய
அயனிகளுடன் இணைவதற்காகச் செல் சவ்வின் வெளிப்பரப்பிற்குச் செல்கிறது (படம் 11.25).
இக்கொள்கை இரு கோட்பாடுகள் வழியாக விளக்கப்படுகிறது.
அ) லூண்டிகார்த் சைட்டோ குரோம் உந்து செயல் கோட்பாடு - (Lundegardh's Cytochrome Pump Theory)
லூண்டிகார்த்
மற்றும் பர்ஸ்ட்ரோம்
(1933) ஆகியோர் சுவாசித்தலுக்கும் எதிர்மின் அயனி உள்ளெடுப்பதற்குமான தொடர்பினைக் கண்டறிந்தனர்.
இதன்படி, ஒரு தாவரத்தினை நீரிலிருந்து உப்பு கரைசலுக்கு மாற்றும்போது சுவாசித்தல் வீதம்
அதிகரிக்கிறது. இதற்கு எதிர்மின் அயனி சுவாசித்தல்
அல்லது உப்பு சுவாசித்தல் என்று பெயர்.
இதனை அடிப்படையாக வைத்துச் சைட்டோகுரோம் உந்து செயல் கொள்கையினை லூண்டிகார்த்
(1950, 1954) கீழ்காணும் அனுமானங்களின் அடிப்படையில் முன்மொழிந்தார்.
1) நேர்மின் அயனியின் உள்ளெடுப்பும் எதிர்மின் அயனியின்
உள்ளெடுப்பு முறையும் வெவ்வேறானவை.
2) எதிர்மின் அயனியானது சைட்டோகுரோம் சங்கிலி வழியாக
ஆற்றல் சார் உள்ளெடுப்பு மூலமாகவும் நேர்மின் அயனியானது ஆற்றல் சாரா உள்ளெடுப்பு மூலமாகவும்
உறிஞ்சப்படுகிறது.
3) செல்சவ்வின் வெளிப்பரப்பில் ஆக்ஸிகரணமும் உட்பரப்பில்
ஒடுக்கமும் நடைபெறுவதால் ஆக்ஸிஜன் சரிவு ஏற்படுகிறது.
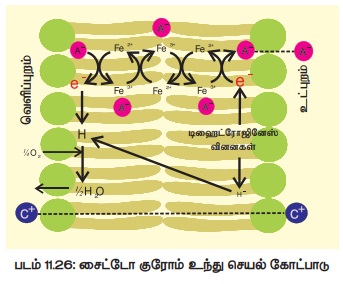
இக்கோட்பாட்டின்படி, சவ்வின் உட்பரப்பில் டிஹைட்ரோஜினேஸ்
நொதியின் செயல்பாடு காரணமாகப் புரோட்டான்களும் (H+) எலெக்ட்ரான்களும்
(e-) உருவாகின்றன. எலெக்ட்ரான்கள் எலெக்ட்ரான் கடத்துச் சங்கிலி வழியாக
வெளிச்செல்லும்போது அதே மின்சுமையினைச் சார்ந்த எதிர்மின் அயனிகள் உள் நுழைகின்றன.
இவ்வாறு உள் நுழையும் எதிர்மின் அயனிகளை ஆக்ஸிஜனேற்றமடைந்த சைட்டோகுரோம் ஆக்ஸிடேஸ் நொதி
ஏற்றுக்கொண்டு சைட்டோகுரோம் சங்கிலியின் அடுத்தடுத்த நிலைக்குக் கடத்துகின்றன (படம்
11.26).
மேலும் இக்கோட்ப்பாட்டின்படி சவ்வின் உட்பரப்பில் எதிர்மின் அயனிகள் (A+) சேகரமாவதான் காரணமாக ஏற்படும் மின்சரிவு வாட்டத்தினை ஈடுசெய்ய நேர்மின் அயனிகள் (C+) ஆற்றல் சாரா முறையில் உள் நுழைகின்றன.
இக்கோட்பாட்டின் குறைகள் பின்வருமாறு,
1) இக்கோட்பாட்டிற்கு மாறாக நேர்மின் அயனிகளும் சுவாசித்தலைத் தூண்டுகின்றன.
2) அயனிகளைத் தேர்வுசெய்து உள்ளெடுப்பதை இக்கொள்கையால் விளக்க இயலவில்லை.
3) இது எதிர்மின் அயனிகளின் உள்ளெடுப்பினை மட்டுமே விளக்குகிறது.
ஆ) பென்னட் கிளார்க்கின் புரத - லெசித்தின் கோட்பாடு :
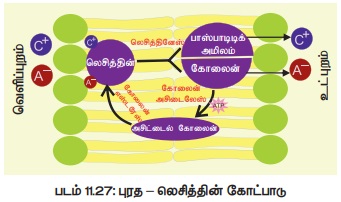
பென்னட் கிளார்க் 1956ல் முன்மொழியப்பட்ட இக்கோட்பாட்டின்படி கடத்தியானது ஒரு புரதமாகும். இது லெசித்தின் எனப்படும் பாஸ்பாடைடுடன் இணைந்து காணப்படுகிறது. இக்கடத்தியானது ஆம்போடெரிக் (அமிலமாகவும் காரமாகவும் செயல்லாற்ற வல்லது) இயல்பு கொண்டதால் நேர் மின் அயனி மற்றும் எதிர்மின் அயனி ஆகிய இரண்டுடனும் சேர்ந்து லெசித்தின் - அயனி கூட்டமைவாக மாறுகிறது. இக்கூட்டமைவு செல்சவ்வின் உட்புறத்தில் பிரிந்து பாஸ்பாடிடிக் அமிலம் மற்றும் கோலைன் ஆகியவற்றுடன் அயனிகளையும் விடுவிக்கிறது. பிரிந்த பாஸ்பாடிக் அமிலம் (Phosphatidic acid) மற்றும் கோலைன் ஆகியவை கோலைன் அசிட்டைலேஸ் மற்றும் கோலைன் எஸ்டரேஸ் எனும் நொதியினால் மீளவும் ஒன்றிணைந்து லெசித்தின் புரதத்தினை உருவாக்குகின்றன (படம் 11.27). இவ்வகை லெசித்தின் உருவாக்கத்திற்கு ATP தேவைப்படுகிறது.
3. டோனன் சமநிலை
சில அயனிகளால் செல்சவ்வின் வழியாக ஒருபோதும் ஊடுருவி வெளியேற முடிவதில்லை. இவை பொறியில் அகப்பட்டது போலச் செல்லுக்குள் உள்ளன. இவற்றுக்கு நிலைத்த அயனிகள் என்று பெயர். இவ்வகை அயனிகள் அதன் மாற்று மின்சுமை கொண்ட அயனிகளால் சமன்படுத்தப்பட வேண்டும். எடுத்துக்காட்டாகச் செல்லினுள் நிலைத்த எதிர்மின் அயனிகள் இருப்பதாகக் கருதினால், இதனைச் சமன்படுத்த நேர்மின் அயனிகள் அதிக அளவில் வழக்கமான அயனிப் பரிமாற்றத்துடன் கூடுதலாக உள்ளெடுக்கப்பட வேண்டும். எனவே செல்லுக்குள் நேர்மின் அயனிகளின் செறிவானது செல்லுக்கு வெளியில் உள்ளதை விட அதிகமாக இருக்க நேரிடுகிறது. மின்னாற்றல் சமன் மற்றும் பரவல் நிகழ்வு ஆகிய இரண்டையும் மின் அயனிகளால் கட்டுப்படுத்தப்படுவதால் இது டோனன் சமநிலை என அழைக்கப்படுகிறது.